In Aplysia
Introduction
Habituation is a response decrement to the repeated application of a given stimulus that cannot be attributed to sensory adaptation, or sensory or motor fatigue (Thompson and Spencer, 1966; Rankin et al., 2009). Although apparently ubiquitous throughout the animal kingdom, and commonly regarded as the simplest form of learning, habituation remains poorly understood with respect to its underlying neuronal mechanisms, especially in vertebrates. Some progress has been made, withal, toward an understanding of the neural basis of habituation in simpler invertebrate organisms, especially the crayfish (Krasne and Teshiba, 1995), the mollusk Aplysia (Glanzman, 2009), and the nematode Caenorhabditis elegans (Giles and Rankin, 2009). Nonetheless, fifty-fifty in these relatively simple organisms, in that location are large gaps in our understanding of the cell biology of habituation.
Habituation tin exhibit both short- and long-term forms. In Aplysia the siphon-elicited gill- and siphon-withdrawal reflex can undergo habituation that persists for several weeks (Carew et al., 1972). This class of long-term habituation (LTH) is characterized by long-term synaptic depression of the sensorimotor pathway that mediates the withdrawal reflex (Carew and Kandel, 1973; Castellucci et al., 1978), as well every bit past retraction of the presynaptic terminals and branches of siphon sensory neurons (Bailey and Chen, 1983, 1988a). Even so, the signaling pathways whose activation during LTH triggers these long-term cellular changes remain fairly obscure. In a previous study (Ezzeddine and Glanzman, 2003) we used a reduced preparation to report the signaling pathways involved in LTH of the gill-withdrawal reflex (GWR) in Aplysia. We found that LTH of the GWR depends on poly peptide synthesis, as well as activation of protein phosphatases 1 and 2A and postsynaptic glutamate receptors, specifically, α-amino-iii-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) and N-methyl-D-aspartate (NMDA) receptors. Here, we used our reduced preparation to exam the roles of RNA synthesis, calcineurin activity and 50-type Ca2+ channel activation in LTH in Aplysia.
Materials and Methods
We have previously described the Materials and Methods in detail (Ezzeddine and Glanzman, 2003). Briefly, adult Aplysia californica (75–150 g) were obtained from a local supplier (Alertness Marine, Redondo Embankment, CA, United states of america) and housed for ≥24 h prior to the start of the experiments. During an experiment the animal was initially anesthetized with isotonic MgCltwo, and then the curtain shelf, gill, siphon, and tail were dissected away from the rest of the trunk, together with the CNS (minus the buccal ganglia), which was left continued to the siphon and gill via the siphon and branchial nerves, respectively. The abdominal artery was cannulated with polyethylene tubing (0.024 in OD, 0.011 in ID, Intramedic, Parsippany, NJ, USA), and this was continued to a peristaltic pump. During experiments the abdominal avenue was perfused (rate = 1.v ml/hr) with aerated normal bogus seawater (ASW, 15°C) via the cannula. The cannula was also used to selectively administrate drugs to the intestinal ganglion. Following cannulation of the abdominal artery, the preparation was pinned to the Sylgard-lined bottom of a Lucite experimental chamber. The siphon was left unpinned. The afferent vein of the gill was cannulated with polyethylene tubing and perfused with chilled, aerated ASW. The cannula in the afferent vein was secured with a surgical silk suture, and the suture was continued to a forcefulness transducer (Model 1040 or 1030, ADInstruments, Grand Junction, CO, USA), which was used to measure the GWR. Habituating and test stimuli were delivered to the siphon via pairs of Teflon-insulated platinum wires (0.005 mm in diameter, #773000, AM Systems, Carlsborg, WA, U.s.). One wire was inserted into each side of the siphon, and a footing wire was placed in the bath. The intensity of the stimuli (500 ms trains [25 Hz] of ten ms current pulses) was set for each preparation to exist only suprathreshold for reliably eliciting gill withdrawal, and remained fixed throughout an experiment. 1 side of the siphon (Trained) received both the test and habituating stimulation, whereas the other side (Untrained) received merely the test stimulation.
After the intensity of the stimuli had been determined for the Trained and Untrained sides, the grooming was rested for 60–xc min. At the terminate of the residue period a single stimulus was delivered to the Trained side of the siphon. Preparations that did not respond to this initial stimulus with a visible gill contraction were discarded. Furthermore, data from a grooming were accepted only if the posttest response of the Untrained side was at to the lowest degree 80% of the pretest response.
All drugs were purchased from Sigma Aldrich (St. Louis, MO, USA). The drugs were initially dissolved in DMSO, and then diluted to their final concentrations in artificial seawater (ASW). (The final concentration of DMSO in the infusate was 0.i–0.two%.) The drugs were directly delivered to the intestinal ganglion via the cannula in the abdominal artery. The irreversible transcriptional inhibitor actinomycin-D was infused into the abdominal ganglion for one h prior to the pretests, and and then washed out. All other drugs were infused into the abdominal ganglion starting immediately after the pretests, and and so washed out of the abdominal ganglion with ASW prior to the posttests. In control preparations ASW, containing the same concentration of DMSO as the infusate used in the matched experimental preparations, was infused into the intestinal artery, and then washed out with normal ASW prior to the posttest. The experimental preparations and the control preparations used to test the effect of an experimental drug on habituation were from animals that were all collected at the same fourth dimension.
The side of the siphon that was called to exist the Trained side was alternated systematically between left and correct sides. The Trained side initially received a pretest stimulus, and v min later a pretest stimulus was delivered to the other side of the siphon (Untrained side). 1 h subsequently the pretest stimulus to the Untrained side, the Trained side received habituation training, which consisted of 5 blocks of stimuli (interblock interval = ninety min). During each block thirty stimuli (ISI = thirty or 60 southward) were delivered to the Trained side. Following a two h rest period after habituation grooming, the Trained and Untrained sides each received a single posttest stimulus. The two posttest stimuli were separated by v min.
The pinnacle aamplitude (maximum–minimum) of each gill contraction was determined using a digital information conquering system (MacLab 4 s/2e or PowerLab 8 s, ADInstruments, Colorado Springs, CO, Us). The minimum was the base line response recorded when the gill was in a relaxed country 1 s prior to stimulation. The pretest value of the GWR was set up to be equal to 100% for the Trained and Untrained sides, and subsequent GWRs were normalized to the pretest value. All responses in the Results are expressed every bit the mean normalized response ± SEM. Not-parametric statistical tests were used for all comparisons. Wilcoxon tests were used for paired comparisons, and Mann–Whitney tests were used for unpaired comparisons unless otherwise indicated. All significance values reported represent two-tailed levels of significance.
Results
Long-Term Habituation Depends on RNA Synthesis
To exam whether LTH of the GWR requires transcription, actinomycin-D (40 μM in ASW with 0.two% DMSO) was infused into the abdominal ganglion. Actinomycin-D's effects are irreversible; therefore, the drug was applied for ane h, and so done out with ASW immediately before the start of the pretests. The actinomycin-D treatment (n = half-dozen) blocked LTH (Figure 1A). The divergence between the pretest and posttest withdrawal in response to stimulation of the Trained side was not pregnant (mean normalized posttest response = 99.three ± 12.v%, p > 0.6). Furthermore, the posttest GWR evoked by stimulation of the Untrained siphon side was not significantly different from the pretest GWR (mean normalized posttest response = 111.ii ± ix.five%, p > 0.1). Finally, there was no statistical difference between the Trained and Untrained posttest GWRs (p > 0.1).
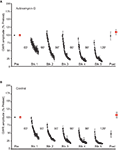
Figure 1. Consequence of actinomycin-D treatment on LTH. (A) Experiments in which preparations were treated with the irreversible transcriptional inhibitor actinomycin-D prior to habituation training (north = 6). The drug blocked the induction of LTH. (B) Data for control experiments (n = vi). Hither, the abdominal ganglion was perfused with normal ASW plus DMSO for the time corresponding to the period of actinomycin-D treatment. The habituation training produced significant LTH of the GWR to stimulation of the Trained side of the siphon. In this and the following figures the solid circles represent the responses to Trained side stimulation, whereas the solid squares represents the responses to Untrained side stimulation.
In the command experiments (north = 6) ASW containing 0.2% DMSO was infused into the abdominal ganglion 1 h before the pretests, and washed out immediately before the start of the experiment. The preparation produced LTH of the GWR to Trained side stimulation (mean normalized posttest response = 47.three ± 10.2%, p < 0.04 for the comparing with the Trained pretest response) (Figure 1B). By contrast, the posttest GWR to stimulation of the Untrained side of the siphon did not differ significantly from the pretest GWR (mean normalized response = 108 ± viii.8%, p > 0.4). Also, the posttest response evoked by Trained side stimulation was significantly less than that evoked past Untrained side stimulation (p < 0.003). In addition, there was a significant deviation betwixt the Trained posttest GWRs of the actinomycin-D-treated and control groups (p < 0.003). The Untrained posttest responses of the ii groups, however, did not differ significantly (p > 0.8). Importantly, despite the fact that actinomycin-D treatment blocked LTH, Trained-site stimulation nonetheless produced normal short-term habituation (STH) of the GWR in the actinomycin-treated preparations. (Compare the Trained responses to the Block 1 habituating stimuli in Figures 1A,B).
To control for not-specific furnishings of actinomycin-D, we tested the effect of a reversible transcriptional inhibitor, 5,six-dichloro-1-β-D-ribofuranosylbenzimidazole (DRB) (Sehgal and Tamm, 1978; Yamaguchi et al., 1998; Apergis-Schoute et al., 2005), on LTH. DRB (150 μM in ASW with 0.2% DMSO) was nowadays in the abdominal ganglion throughout the training menstruation. The presence of DRB during habituation training blocked LTH (n = 9, Effigy 2A). At that place was no significant alter in the Trained GWR (hateful normalized postest response = 115.1 ± 10.9%, p > 0.2 for the comparison with the pretest response). The GWR to stimulation of the Untrained side also did not change (hateful normalized Untrained posttest response = 122.6 ± xix.one%, p > 0.seven for the comparison with the pretest response). In addition, the Trained and Untrained posttests did not differ (p > 0.eight). Control experiments, in which ASW with 0.ii% DMSO was infused into the intestinal artery, were performed using the aforementioned protocol as the DRB experiments. The habituation training produced pregnant LTH of the Trained GWR in the control experiments (n = ix, Effigy 2B). There was a significant decrease in posttest response to Trained side stimulation (mean normalized posttest response = 75.vi ± 8.eight% p < 0.04 for the comparing with the pretest response). Yet, there was a meaning increase in the GWR to stimulation of the Untrained side (mean normalized posttest response = 118.4 ± 7.5%, p < 0.03 for the comparing with the Untrained pretest response). Furthermore, the posttest Untrained GWR was significantly greater than the posttest Trained GWR (p < 0.004). Finally, the posttest GWRs to Trained side stimulation differed significantly in the DRB-treated and control preparations (p < 0.02).
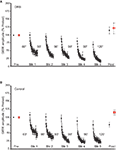
Figure 2. Issue of DRB treatment on LTH. (A) Information from experiments (n = ix) in which preparations were treated with the reversible transcriptional inhibitor DRB during habituation training. DRB infusion disrupted LTH. (B) Data for the command experiments (n = 9). Hither, training was carried out in the presence of ASW plus DMSO, which produced LTH on the Trained side and sensitization on the Untrained side.
The effect of the habituation training on the GWR to Untrained side stimulation in the control experiments suggests that the training produced some generalized sensitization, as well as pathway-specific LTH, in these experiments. In our earlier report (Ezzeddine and Glanzman, 2003) we also observed that the GWR to Untrained side stimulation sometimes exhibited sensitization after habituation training, and that there was often sensitization of the GWR to Trained side stimulation during training in the presence of drugs that blocked LTH. We believe, every bit discussed in that paper, that these phenomena reflect the fact that the habituating stimuli can activate both habituating and sensitizing neuronal processes, as originally proposed by Groves and Thompson (1970). According to Groves and Thompson, the ultimate behavioral outcome, whether habituation, sensitization, or no change, is adamant past the relative strengths of these two countervailing processes. It should be noted, yet, that both the sensitization of the GWR on the Untrained side, and the sensitization of the reflex on the Trained side during habituation training in the presence of inhibitors of LTH (e.1000., Blocks 2 and three, Figure 1A), were highly variable and non always observed.
Long-Term Habituation Requires Calcineurin Activity
We previously constitute that LTH depends on activation of NMDA-type receptors in abdominal ganglion. Activation of postsynaptic NMDA receptors would be expected to produce a rise in intracellular Ca2+ in neurons of the GWR excursion. The Ca2+/calmodulin-dependent phosphatase calcineurin has been shown to be involved in both long-term synaptic low (Mulkey et al., 1994) and memory extinction (Baumgartel et al., 2008) in mammals. Given that long-term depression (LTD) of the sensorimotor synapse (Lin and Glanzman, 1996) is a candidate mechanism for LTH in Aplysia (see Ezzeddine and Glanzman, 2003; Glanzman, 2009), nosotros wished to know whether LTH was regulated by calcineurin activity. Accordingly, nosotros tested the result of ascomycin (10 μM in ASW with 0.2% DMSO), a selective calcineurin inhibitor (Sierra-Paredes and Sierra-Marcuño, 2008), on LTH. Ascomycin application during habituation training blocked LTH (n = 7, Effigy 3A). The difference between the pretest and posttest responses for Trained side stimulation was non pregnant (hateful normalized posttest response = 104.2 ± xiv%, p > 0.nine). However, as was the example for the command experiments in Effigy 2B, the posttest response for the Untrained side exhibited significant enhancement (mean normalized posttest response = 143.ii ± xv.7%, p < 0.05 for the comparison with the pretest response). The difference between the posttest responses for the Trained and the Untrained sides, withal, was non significant (p > 0.ane). Control experiments (n = 5) were performed with rapamycin (ten μM in ASW with 0.2% DMSO) present in the cannula during training (Figure 3B). Rapamycin has the aforementioned binding site on calcineurin as ascomycin, but rapamycin does non inhibit dephosphorylation by calcineurin (Schreiber and Crabtree, 1992). In contrast to the consequence of ascomycin, rapamycin did not impact LTH. Thus, habituation training resulted in a significantly reduced GWR for the Trained side (mean normalized posttest response = 51.4 ± 13%, p < 0.03 [t-test used here for the comparing with the pretest response]). The deviation between the pretest and posttests responses for Untrained side, however, was not significant (mean normalized posttest GWR = 93.4 ± vi.7%, p > 0.three). As would be expected from these results, the posttest responses for the Trained and Untrained sides differed significantly in the experiments involving rapamycin treatment (p < 0.03). Finally, the posttest GWR to Trained side stimulation was significantly less in the rapamycin-treated preparations than in the ascomycin-treated preparations (p < 0.05). Find that, although blockade of calcineurin activeness with ascomycin disrupted LTH, it did not affect STH (Figure 3A).
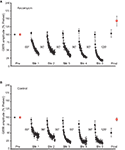
Figure 3. Event of inhibition of calcineurin on LTH. (A) Data from experiments (n = 7) in which the intestinal ganglion was infused with ascomycin during habituation grooming. Inhibition of calcineurin blocked LTH. (B) Control data from experiments (n = 5) in which a control drug, rapamycin, which does not inhibit protein phosphatases, was present in the abdominal ganglion during training. Training in the presence of rapamycin produced LTH of the GWR to Trained side stimulation.
Activation of L-Type Voltage-Gated Catwo+ Channels Appears to be Necessary for LTH
Nosotros previously found that activation of AMPA receptors, also as of NMDA receptors, contributes to LTH of the GWR (Ezzeddine and Glanzman, 2003). It is therefore possible that depolarization-induced activation of L-type voltage-gated Ca2+ channels (LVGCCs) is upstream of calcineurin activity during LTH. To test this possibility, we examined the effect of blockade of LVGCCs on LTH. Nosotros infused nitrendipine (100 μM in ASW with 0.14% DMSO), an antagonist of LVGCCs (Bolshakov and Siegelbaum, 1994), into the abdominal ganglion during habituation grooming (n = half dozen). Nitrendipine treatment blocked LTH to stimulation of the Trained side of the siphon (mean normalized posttest response = 116.8 ± xiii%, p > 0.3 for the comparison with the pretest response) (Figure 4A). In these experiments habituation training did not alter the GWR to stimulation of the Untrained side; the mean normalized Untrained posttest response (157.viii ± 39.iv%, was not significantly unlike from the hateful normalized Untrained pretest response (p > 0.iv), although the variability in the Untrained responses was absolutely large. Furthermore, the GWR evoked by posttest stimulation of the Trained side was not significantly different from that evoked by posttest stimulation of the Untrained side (p > 0.9). In control experiments (n = 8), performed at the aforementioned time as the nitrendipine experiments, the intestinal ganglion was infused ASW plus 0.14% DMSO. Here, habituation training produced significant LTH of the GWR to stimulation of the Trained side (hateful normalized posttest response = 55.1 ± 12.4%, p < 0.02 for the comparing with the pretest response) (Figure 4B). At that place was no significant alter in the GWR to stimulation of the Untrained side (mean Untrained normalized posttest response = 124.4 ± 15.3%, p > 0.1 for the comparison with the Untrained pretest response). Furthermore, the Trained side posttest GWR and the Untrained side posttest GWR differed significantly (p < 0.004). Finally, the departure between the posttest response to the Trained side stimulation in the nitrendipine group and that in the control grouping was highly significant (p < 0.005). These results back up the idea that LVGCC activity is essential for LTH, although such action appears not to be necessary for STH (Effigy 4A).
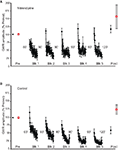
Effigy 4. Outcome of inhibiting LVGCCs on LTH. (A) Data from experiments (due north = 6) in which nitrendipine was nowadays during habituation training. The drug blocked the induction of LTH. (B) Data from control experiments (north = eight) in which grooming occurred in the presence of ASW plus DMSO. The training resulted in LTH of the GWR.
Discussion
Office of Gene Transcription in LTH in Aplysia
Retentiveness in Aplysia has been divided into 3 stages: brusk-term, intermediate-term and long-term (Goelet et al., 1986; Sutton and Carew, 2002). These 3 stages are distinguished by both their temporal and mechanistic properties. Temporally – although their fourth dimension courses overlap somewhat – short-term memory (STM) lasts from seconds to ∼30 min; intermediate-term retentiveness (ITM) lasts from ∼thirty min to 3 h; and long-term memory lasts for >10 h. Mechanistically, STM depends exclusively on posttranslational changes; ITM requires protein synthesis, but non gene transcription; and LTM requires both translation and transcription. Even so, most of the previous work characterizing the mechanisms underlying the different phases of retention in Aplysia has concerned but one type of retentivity – sensitization of the withdrawal reflex. Although an early study demonstrated that a neural correlate of STH of the GWR did not require poly peptide synthesis (Schwartz et al., 1971), there has been piffling attempt until recently to make up one's mind whether there are unlike phases of retentivity for habituation that correspond mechanistically to those for sensitization. We previously showed that the habituation of the GWR that persisted for 1–6 h following a 5 h period of spaced training required protein synthesis (Ezzeddine and Glanzman, 2003). But in that report we did non examination whether this form of habituation retentivity also required gene transcription. Here, we have shown that the habituation resulting from a similar grooming protocol requires transcription, considering it was blocked by actinomycin-D and DRB. Therefore, this form of habituation fits the definition of LTM as it has commonly been defined in Aplysia (Goelet et al., 1986). It will exist interesting in the future to determine whether there is a preparation protocol that produces an intermediate form of habituation memory, one that requires translation, but not transcription.
Homosynaptic Depression Cannot Account for LTH in Aplysia
Information technology has long been believed that the mechanism of habituation of the gill- and siphon-withdrawal reflex is due exclusively to presynaptic changes. This mechanism, homosynaptic depression (HSD), is thought to involve decreased transmitter release from presynaptic terminals due to repeated activation of sensory neurons (Castellucci and Kandel, 1974; Armitage and Siegelbaum, 1998). Currently, the mechanism underlying HSD in Aplysia is controversial. It was originally proposed that HSD was acquired past depletion of the readily releasable pool of presynaptic vesicles (Gingrich and Byrne, 1985; Hochner et al., 1986; Bailey and Chen, 1988b). More than recent show, however, indicates that HSD results from all-or-none silencing of presynaptic release sites (Gover and Abrams, 2009).
Although HSD may indeed be a major mechanism of STH, the data from this and our previous written report (Ezzeddine and Glanzman, 2003) argue strongly confronting the notion that LTH can be explained past exclusively presynaptic changes. First, HSD does not depend on glutamate receptor activity (Armitage and Siegelbaum, 1998), whereas LTH does (Ezzeddine and Glanzman, 2003). Second, LTH requires activation of LVGCCs (present written report), which is most likely acquired by AMPA receptor-mediated postsynaptic depolarization (see Ezzeddine and Glanzman, 2003). Studies by Bailey and Chen (1983, 1988a) indicate that LTH is accompanied past presynaptic morphological changes, including fewer vesicles in sensory neuron-associated agile zones, and a subtract in the number of varicosities per sensory neuron. These morphological data, together with our data, support the conclusion that LTH involves both pre- and postsynaptic long-term cellular changes. If correct, this conclusion raises the intriguing possibility that the pre- and postsynaptic changes are coordinated past transsynaptic signals (see Glanzman, 2009, for further discussion).
Role of Calcineurin and LVGCCs in LTH
The data from the present study suggest that increased intracellular Ca2+ plays a key role in LTH. One probable source of elevated intracellular Ca2+ is influx through postsynaptic NMDA receptors (Ezzeddine and Glanzman, 2003). Activated LVGCCs, due to AMPA receptor-mediated postsynaptic depolarization, correspond another potential source of the elevated intracellular Ca2+ necessary for LTH. Our finding that ascomycin blocks the induction of LTH indicates that calcineurin is one of the proteins whose activity is stimulated past increased intracellular Ca2+. Interestingly, calcineurin activeness has been implicated in both LTD (Mulkey et al., 1994) and depotentiation (Jouvenceau et al., 2003) in the mammalian brain, as well every bit in some forms of extinction in mammals (Lin et al., 2003; Baumgartel et al., 2008). Nosotros practice not yet know what downstream pathways are stimulated past calcineurin during LTH. One action linked with calcineurin activity in the mammalian brain is endocytosis of AMPA receptors, which is believed to be a key event in LTD (Beattie et al., 2000). Endocytosis of AMPA-type receptors could well exist ane consequence of calcineurin activity during habituation of the GWR (see Glanzman, 2009), because exocytosis of AMPA-type receptors has previously been shown to play a role in synaptic facilitation and enhancement of the withdrawal reflex in Aplysia (Chitwood et al., 2001; Li et al., 2005, 2009).
Potential Office of LTD in LTH in Aplysia
The synaptic mechanisms that underlie LTH of the withdrawal reflex remain to exist identified. One potential mechanism that is consistent with the information in this and our previous study (Ezzeddine and Glanzman, 2003) is LTD induced by elevated postsynaptic Ca2+ (Malenka and Carry, 2004). A form of LTD of the sensorimotor synapse that depends partly on elevated postsynaptic Ca2+ has been demonstrated in dissociated cell culture (Lin and Glanzman, 1996); just this form of synaptic low has not yet been experimentally linked to LTH. A problem for the idea that activity-dependent LTD is a mechanism of LTH is that induction of activity-dependent LTD requires 1 Hz electrical stimulation, which is quite different from the rate of behavioral stimulation that results in LTH. Further complicating the issue of the synaptic machinery of LTH is the demonstration past Montarolo et al. (1988) that homosynaptic activation of the sensorimotor synapse with a charge per unit of stimulation designed to mimic the synaptic stimulation that occurs during LTH training fails to induce long-term (24 h) depression of the synapse. The credible failure of naturalistic homosynaptic action to induce LTD of the sensorimotor synapse suggests that homosynaptic activation of the sensorimotor pathway, past itself, may exist insufficient for LTH in Aplysia. The possibility that LTH may depend, at least in part, on one or more heterosynaptic pathways is supported by the finding that spaced applications of the endogenous inhibitory neuropeptide, FMRFamide, can produce low of the sensorimotor synapse that persists for ≥24 h (Montarolo et al., 1988). Both heterosynaptic, inhibitory (GABAergic), and homosynaptic pathways take been shown to play roles in habituation of the crayfish tailflip escape response (Krasne and Teshiba, 1995).
Mechanistic Similarities between Habituation and Extinction
Habituation, like extinction, is the waning of a response due to repeated presentation of an unchanging stimulus. The major phenomenological stardom between habituation and extinction is that habituation is the waning of an innate response, whereas extinction is the waning of a conditioned response. Despite this distinction, the behavioral similarity between habituation and extinction has led to the speculation that these 2 forms of memory may share common mechanisms (McSweeney and Swindell, 2002). Although controversial, there is experimental support for this thought (Kamprath et al., 2006). We have found that LTH in Aplysia requires the action of calcineurin and LVGCCs. Both of these processes have also been implicated in extinction (Lin et al., 2003; Barad et al., 2004; Suzuki et al., 2004, 2008; Cain et al., 2005; Baumgartel et al., 2008) (simply run into Schafe, 2008). Maybe, therefore, an understanding of the cellular mechanisms that mediate LTH of the GWR in Aplysia will contribute important mechanistic insights into the considerably more circuitous phenomenon of extinction.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of whatever commercial or financial relationships that could be construed as a potential conflict of involvement.
Acknowledgments
We give thanks A. Roberts and D. Cai for helpful comments on the manuscript. This work was supported by grants R01 MH068543 and R37 NS029563 from the United states National Institutes of Health, and a grant from the Sprint Foundation (David L. Glanzman).
References
Apergis-Schoute, A. G., Debiec, J., Doyere, V., LeDoux, J. E., and Schafe, G. Eastward. (2005). Auditory fear conditioning and long-term potentiation in the lateral amygdala require ERK/MAP kinase signaling in the auditory thalamus: a role for presynaptic plasticity in the fear system. J. Neurosci. 25, 5730–5739.
Pubmed Abstract | Pubmed Full Text | CrossRef Total Text
Armitage, B. A., and Siegelbaum, S. A. (1998). Presynaptic consecration and expression of homosynaptic low at Aplysia sensorimotor neuron synapses. J. Neurosci. 18, 8770–8779.
Pubmed Abstract | Pubmed Full Text
Bailey, C. H., and Chen, M. (1988a). Long–term memory in Aplysia modulates the total number of varicosities of single identified sensory neurons. Proc. Natl. Acad. Sci. U.S.A. 85, 2373–2377.
CrossRef Total Text
Bailey, C. H., and Chen, M. (1988b). Morphological basis of short-term habituation in Aplysia. J. Neurosci. 8, 2452–2459.
Barad, Thou., Blouin, A. M., and Cain, C. K. (2004). Like extinction, latent inhibition of conditioned fearfulness in mice is blocked past systemic inhibition of fifty-type voltage-gated calcium channels. Acquire. Mem. 11, 536–539.
Pubmed Abstruse | Pubmed Full Text | CrossRef Full Text
Baumgartel, Thousand., Genoux, D., Welzl, H., Tweedie-Cullen, R. Y., Koshibu, K., Livingstone-Zatchej, Yard., Mamie, C., and Mansuy, I. M. (2008). Control of the institution of aversive retentiveness by calcineurin and Zif268. Nat. Neurosci. 11, 572–578.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Beattie, E. C., Carroll, R. C., Yu, X., Morishita, W., Yasuda, H., von Zastrow, Grand., and Malenka, R. C. (2000). Regulation of AMPA receptor endocytosis by a signaling mechanism shared with LTD. Nat. Neurosci. 3, 1291–1300.
Pubmed Abstruse | Pubmed Full Text | CrossRef Full Text
Cain, C. M., Godsil, B. P., Jami, S., and Barad, M. (2005). The l-type calcium channel blocker nifedipine impairs extinction, but not reduced contingency effects, in mice. Learn. Mem. 12, 277–284.
Pubmed Abstruse | Pubmed Full Text | CrossRef Full Text
Castellucci, V. F., and Kandel, E. R. (1974). A quantal analysis of the synaptic depression underlying habituation of the gill-withdrawal reflex in Aplysia. Proc. Natl. Acad. Sci. U.South.A. 71, 5004–5008.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Ezzeddine, Y., and Glanzman, D. 50. (2003). Prolonged habituation of the gill-withdrawal reflex in Aplysia depends on protein synthesis, protein phosphatase activity, and postsynaptic glutamate receptors. J. Neurosci. 23, 9585–9594.
Pubmed Abstract | Pubmed Total Text
Gingrich, M. J., and Byrne, J. H. (1985). Simulation of synaptic low, posttetanic potentiation, and presynaptic facilitation of synaptic potentials from sensory neurons mediating gill-withdrawal reflex in Aplysia. J. Neurophysiol. 53, 652–669.
Pubmed Abstract | Pubmed Full Text
Hochner, B., Klein, Thou., Schacher, Due south., and Kandel, E. R. (1986). Boosted component in the cellular mechanism of presynaptic facilitation contributes to behavioral dishabituation in Aplysia. Proc. Natl. Acad. Sci. UsaA. 83, 8794–8798.
Pubmed Abstract | Pubmed Full Text | CrossRef Total Text
Jouvenceau, A., Billard, J.-One thousand., Haditsch, U., Mansuy, I. Grand., and Dutar, P. (2003). Different phosphatase-dependent mechanisms mediate long-term low and depotentiation of long-term potentiation in mouse hippocampal CA1 area. Eur. J. Neurosci. eighteen, 1279–1285.
Pubmed Abstruse | Pubmed Total Text | CrossRef Full Text
Kamprath, K., Marsicano, One thousand., Tang, J., Monory, Thousand., Bisogno, T., Marzo, V. D., Lutz, B., and Wotjak, C. T. (2006). Cannabinoid CB1 receptor mediates fear extinction via habituation-similar processes. J. Neurosci. 26, 6677–6686.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Krasne, F. B., and Teshiba, T. Chiliad. (1995). Habituation of an invertebrate escape reflex due to modulation by higher centers rather than local events. Proc. Natl. Acad. Sci. U.South.A. 92, 3362–3366.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Li, H. Fifty., Huang, B. Southward., Vishwasrao, H., Sutedja, N., Chen, W., Jin, I., Hawkins, R. D., Bailey, C. H., and Kandel, E. R. (2009). Dscam mediates remodeling of glutamate receptors in Aplysia during de novo and learning-related synapse formation. Neuron 61, 527–540.
Pubmed Abstruse | Pubmed Full Text | CrossRef Full Text
Li, Q., Roberts, A. C., and Glanzman, D. L. (2005). Synaptic facilitation and behavioral dishabituation in Aplysia: dependence upon release of Catwo+ from postsynaptic intracellular stores, postsynaptic exocytosis and modulation of postsynaptic AMPA receptor efficacy. J. Neurosci. 25, 5623–5637.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Lin, C.-H., Yeh, S.-H., Leu, T.-H., Chang, W.-C., Wang, S.-T., and Gean, P.-Westward. (2003). Identification of calcineurin as a key bespeak in the extinction of fright memory. J. Neurosci. 23, 1574–1579.
Pubmed Abstract | Pubmed Full Text
Lin, Ten. Y., and Glanzman, D. Fifty. (1996). Long-term depression of Aplysia sensorimotor synapses in prison cell culture: inductive role of a ascension in postsynaptic calcium. J. Neurophysiol. 76, 2111–2114.
Pubmed Abstruse | Pubmed Full Text
Rankin, C. H., Abrams, T., Barry, R. J., Bhatnagar, S., Clayton, D. F., Colombo, J., Coppola, G., Geyer, Chiliad. A., Glanzman, D. L., Marsland, Southward., McSweeney, F. K., Wilson, D. A., Wu, C. F., and Thompson, R. F. (2009). Habituation revisited: an updated and revised description of the behavioral characteristics of habituation. Neurobiol. Learn. Mem. 92, 135–138.
Pubmed Abstract | Pubmed Total Text | CrossRef Full Text
Schwartz, J. H., Castellucci, V. F., and Kandel, Due east. R. (1971). Functioning of identified neurons and synapses in abdominal ganglion of Aplysia in absence of protein synthesis. J. Neurophysiol. 34, 939–953.
Pubmed Abstruse | Pubmed Full Text
Sutton, M. A., and Carew, T. J. (2002). Behavioral, cellular, and molecular analysis of memory in Aplysia I: intermediate-term retentiveness. Integr. Comp. Biol. 42, 725–735.
CrossRef Full Text
Suzuki, A., Josselyn, S. A., Frankland, P. W., Masushige, Southward., Silva, A. J., and Kida, S. (2004). Retention reconsolidation and extinction have distinct temporal and biochemical signatures. J. Neurosci. 24, 4787–4795.
Pubmed Abstract | Pubmed Full Text | CrossRef Total Text
Suzuki, A., Mukawa, T., Tsukagoshi, A., Frankland, P. Westward., and Kida, Due south. (2008). Activation of LVGCCs and CB1 receptors required for destabilization of reactivated contextual fear memories. Learn. Mem. 15, 426–433.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Source: https://www.frontiersin.org/articles/10.3389/fnbeh.2010.00181/full
0 Response to "In Aplysia"
Post a Comment